
Do Anthrobots Dream of Electric Lungs?
Why anthrobots and archetypes are the same kind of thing

When Philip K. Dick’s novel asked: Do Androids Dream of Electric Sheep?--it was about whether a synthetic being, built without the embedding of a natural lineage, could nonetheless harbor something like longing for the whole it was excluded from. The premise assumes that biological beings carry their wholes inside them in ways that cannot be fully articulated. A dream is one of the places this becomes visible.
Anthrobots invert the question.

If you haven’t come across them yet: anthrobots are self-motile multicellular constructs built from dissociated adult human tracheal epithelial cells. Take a bit of airway lining, separate the cells, put them in permissive conditions, and they self-assemble into motile little forms that swim around and even repair damaged neural tissue across a gap. Freed from the body that organized them, cells that used to help you breathe become something else with a novel behavioral catalog.
Where PKD's synthetic being starts without a lineage embedding, an anthrobot is a biological fragment that had its embedding stripped away.
Their behavior, their morphology, and their bioelectric state are still being shaped by a body that no longer exists. These creatures reveal a synthesis that should feel obvious but doesn't. The claim is simple: parts carry wholes.
"Parts carry wholes" is a claim about a specific kind of information: information about the whole, distributed across many parts, held by each rather than localized in any one.
That’s a working thesis, and I’ve become excited by anthrobots because they reveal what could be a substrate on which to test a novel structural formalism.
Somatic cells carry target morphologies in their bioelectric and epigenetic states. Lineages carry the accumulated developmental learning of their evolutionary history. And--most controversially--psyches carry what Jung called archetypes: structural tendencies inherited by every individual and expressed most clearly when waking constraints loosen.
Anthrobots might be exactly what happens when the constraint comes off.

Let me back up on “target morphology,” because it’s a load-bearing concept here at the somatic scale and it isn’t what most biologists got in school.
The traditional story is that the genome is a blueprint and morphogenesis is its execution. The tissue reads the plans, cells go to their assigned positions and the organism comes out the correct shape. This is wrong in an interesting way. The genome is better described as a component of the hardware on which a distributed computation runs. The shape the tissue converges on is a property of that computation, which is a property of the collective, rather than of any instruction set a single cell is executing.
Michael Levin’s lab has been making this legible for years. Planaria cut into fragments regenerate correctly because each fragment retains access to a landscape whose attractors are “head” and “tail” in the right places. Xenopus embryos with rearranged facial organ positions re-sort toward the canonical face during development. The target is a dynamical feature, a set of attractors in the morphogenetic state space, and it’s robust to perturbations that would break a blueprint.
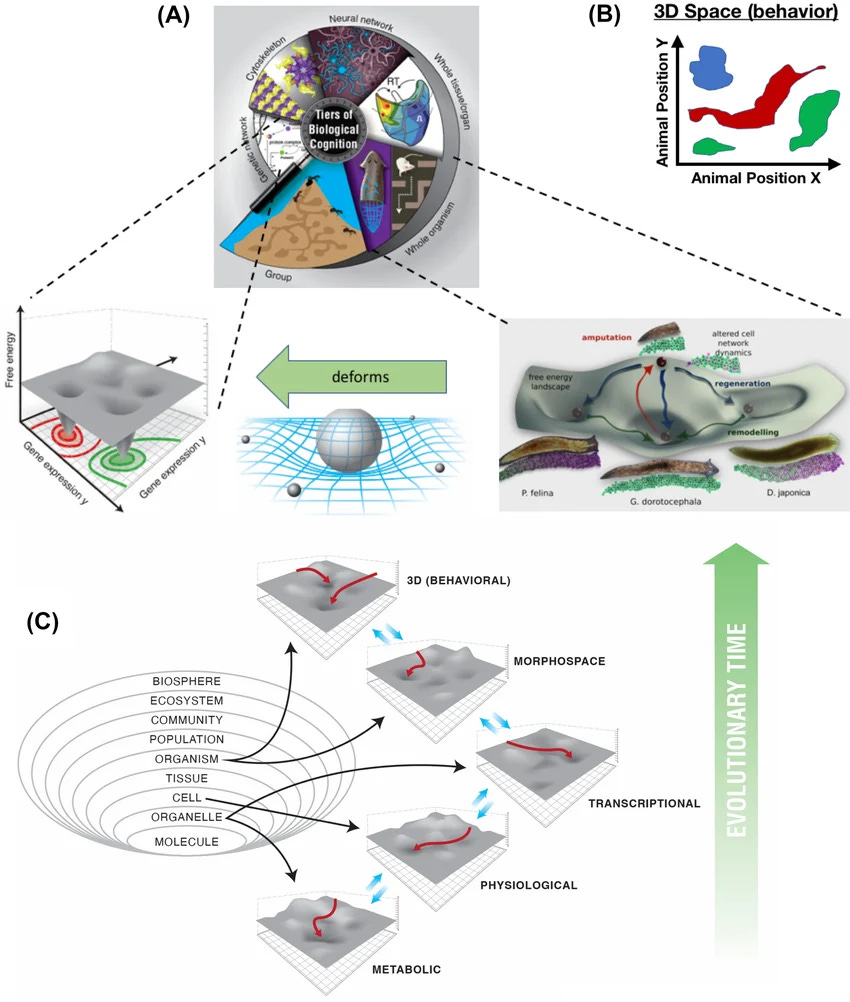
Furthermore, you can edit the landscape without touching the genome. Perturb the bioelectric pattern of a planarian during regeneration, and you can get it to regenerate two heads. Cut that two-headed planarian again, and it regenerates two heads again, stably, despite an unchanged genetic sequence. You’ve rewritten the target without editing the code conventionally assumed to specify it.
This reframes where developmental information actually lives. It’s distributed across membrane potentials, ion channels, gap-junctions, chromatin states, and other biophysical state regulators. It’s editable by interventions that act on any of those. When you take a somatic cell out of its tissue context, it does not lose the landscape. The landscape is partly in the cell, and the cell carries it.
So when you dissociate tracheal epithelium, the cells you end up with are loaded with a bioelectric and epigenetic state shaped by their former position in the airway. Put them in permissive conditions, and they do not rebuild the trachea, but what they do build is a continuation of that landscape rather than a random reorganization. Characteristics like ciliation polarity, motility, and the scale at which they assemble all trace back to what the cells used to be.
The anthrobot is running new software on hardware shaped by the old software. Even after the old software stops executing, it shapes the basin the new dynamics settle into.
The puzzle here is that if the genome→morphology view is wrong, where does the landscape come from?
evolutionary connectionism
Target morphologies are species-typical, stable under perturbation, and precisely adapted to functional requirements. You don’t get that kind of structure out of nothing. So the question is whether evolution is the right kind of process to put it there.
Richard Watson’s work on evolutionary connectionism is the cleanest answer I’ve read. The short version is that natural selection has the formal properties of a learning algorithm. And he really means this formally, not as an analogy. The inductive biases of selection are mathematically equivalent to the inductive biases of connectionist learning. Evolution generalizes over regularities. It compresses repeated structure and deposits priors into the substrate it acts on.
This gives the morphogenetic landscape a clean place of origin: they are the accumulated priors of a learning process that has been running for the full duration of the lineage. The shape the tissue is trying to become is the shape that cumulative selection has taught the substrate to converge on.
That’s one half of the story, but there's another half this framework can't explain on its own.
When Levin and his group started characterizing what anthrobots actually do once dissociated, they found capabilities with no selection history to point at. Anthrobots bridge damaged neural tissue across a gap. Xenobots respond to sound. Anthrobots express transcriptomes roughly half differentially expressed relative to source tissue, including ancient genes and embryonic body-axis genes that have no business being on in adult tracheal epithelium. Neither organism has ever existed as a lifeform under selection pressure. Selection can shape a frog, but there is no “anthrobot ancestor” whose survival depended on wound-healing-across-a-gap.
So where is that capability coming from?
Levin’s answer, which he’s been sharpening publicly over the last year, is that the space of possible forms and behaviors is a structured possibility space with pattern-features that any sufficiently capable substrate can reach into. Evolution discovers useful patterns in that space and exploits them, but the patterns themselves were not deposited by the selection process that found them. A voltage-gated ion channel is, among other things, a transistor, and so once you have transistors, the whole of logic-gate behavior follows for free, without anyone having had to evolve the logic-gate math.
If Levin is right, then the priors-from-selection story is incomplete, and anthrobots make the incompleteness visible. Watson’s framework explains why tracheal cells behave tracheal-ly. It does not explain why they heal neural wounds.
Watson and Levin are not making incompatible claims.
affordances and priors
Watson is describing what selection has fixed into the substrate of a specific lineage--the priors in the learning-theory sense. Levin is describing the broader access that lineage-specific priors haven't yet committed to a single trajectory. The priors are the particular paths selection has worn deep. The possibility space (Levin calls it Platonic; I’ll stay operational and call it the substrate’s affordance set) is the terrain those paths run across.
A somatic cell, on this reading, carries two overlapping things. The first is the priors its lineage has selected for: the bioelectric, epigenetic, and regulatory settings that bias it toward the species-typical form. The second is the substrate-level access to a broader affordance set from which those priors were drawn in the first place: the ion channel repertoire, the gap-junction topology, and the regulatory logic that make that access possible at all.
Under normal embedding, the priors dominate. The cell is tightly constrained by its neighbors and its current function; its trajectory is the one selection has worn into its lineage. Under dissociation, the priors still bias things--the cell is not blank, and the anthrobot’s form is legibly continuous with its tissue of origin. But the broader substrate-level access opens up too, and the cells can find basins they have never occupied before.
So there’s the reconciliation. Both halves are doing work in the anthrobot case. The priors story explains why anthrobots from tracheal epithelium come out legibly tracheal-derived. The affordance-set story explains why they do things no one’s ancestor ever did.
Memory at the morphogenetic scale is literal. Learned structure is stored in a dynamical medium, where the storage medium is also the execution medium.
But the medium is also an interface to a structured possibility space that isn’t itself stored learning. It’s just there, reachable when the constraints that were holding the trajectory in place come off.
Dissociation is decompression. It releases the priors and opens the substrate’s access to what the priors hadn’t fixed a path through.
collective unconscious
I know up to this point I’ve been doing respectable biology and now I’m going to invoke Jung.
Hear me out.
The collective unconscious has a poor reputation in empirical circles, and it deserves some of that reputation, because the weakest versions of the theory (inherited specific images, Lamarckian memories of the mother goddess) are indefensible. Jung got blamed for those versions even when he wasn’t making them. What he actually claimed, in his mature writing, is structurally different, and worth re-reading now that we have better tools to explore the spaces he was describing.
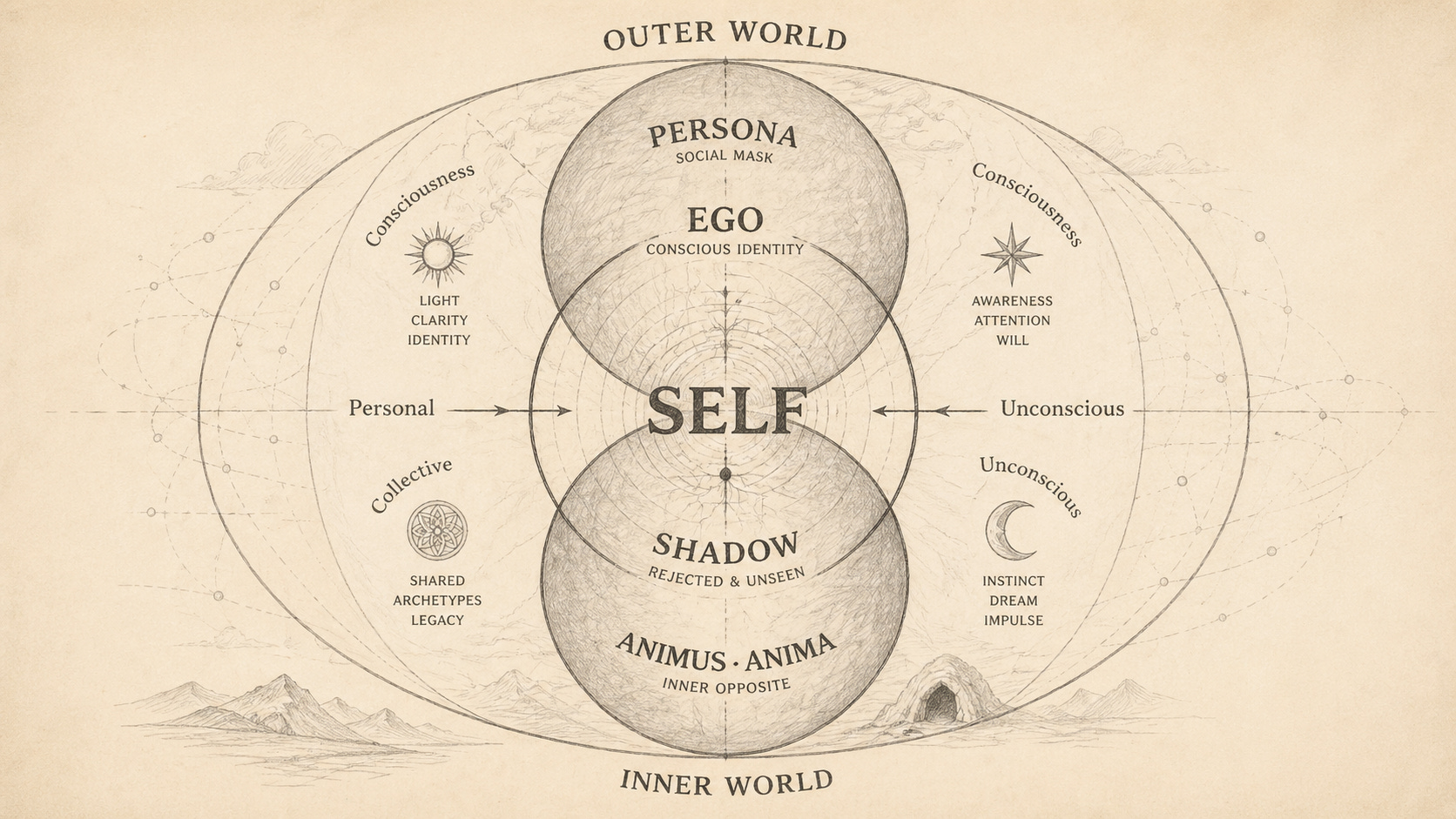
The archetypes, in Jung's careful statement, are formative tendencies: structural dispositions of the psyche to organize experience along particular lines. Read “archetype” and you probably picture a stored image. Jung was explicit that he wasn’t claiming that. The anima/animus, the persona, the shadow, and the self are attractors. When a psyche develops normally, specific contents reliably fall into their basins. What is inherited is the landscape, not the images. The images a given person produces are trajectories through that landscape, shaped by individual history and current constraints.
Everything I've said up to here lands on this next point.
Take the morphogenetic picture and list its formal features. The information is distributed across the cells, not stored in any one of them. It biases a dynamical system toward attractor states rather than toward a fixed endpoint. Those attractor states are species-typical, because they have been shaped by a process operating at the scale of the lineage. The expression of any given attractor at any given moment depends on local substrate state and current constraint: the cell’s epigenetic settings, what neighbors are signaling, what the tissue context is demanding. Perturb the substrate and you can shift which attractor the system finds. Leave it alone and, under canonical conditions, the system returns to the same basin it started from.
Now the psychic picture, with the same list. The information is distributed, not in any one neuron or region, not even in any one episode of experience. It biases a dynamical system (the psyche over time) toward attractor states, which in this domain are the archetypal patterns Jung spent decades cataloguing. Those attractors are species-typical, because they have been shaped by whatever process operates at the scale of the human lineage over its evolutionary history. Expression at any given moment depends on local substrate state (the individual’s history, neurochemistry, developmental trajectory) and current constraint (goal-directed waking attention, social context, sleep versus waking). Perturb the substrate (trauma, drugs, developmental disruption), and you shift which attractor surfaces. Leave it alone and, under canonical conditions, the psyche returns to the same basins.
Five features hold in both cases. The information is distributed rather than stored in any one place. The dynamics are attractor-based rather than endpoint-driven. The attractors are species-typical, because whatever shaped them was operating at lineage scale. Expression on any given day depends on substrate state and on what the context is calling for. And while perturbation can shift the attractors, canonical conditions bring the system back.
If you find the parallelism suggestive but want to stay on the side of caution, you can read it as an analogy: morphogenetic systems and psychic systems happen to share these features, which is interesting but tells us nothing deeper.
The stronger reading is that the five features aren't a coincidence between two domains. They're what you see whenever a dynamical system's trajectory is shaped by information stored across the system's parts, about a pattern bigger than any single part, laid down by a process operating at a slower timescale than the system itself. That formal structure is general. Morphogenesis is one instance. Psychic development is another. Neither is a metaphor for the other. Both are instances of the same thing.
Jung spent decades working out which conditions surface which psychic attractors. He documented how compensation operates when the conscious attitude diverges from the landscape. He catalogued what the signatures of each basin look like in dream, symptom, and symbol.
All of it is data gathered by someone paying close attention to a system we now have better reason to believe actually exists.
And if we take it to exist, the Jungian vocabulary becomes usable the way any technical vocabulary is usable: not as metaphor, but as a record of how a specific instance of the structure behaves.
If you’ve read my recent post on dreams, this is where those threads tie in.
the landscape
My brief claim there was that dreams aren’t noise, replay, or regularization. They are the highest-leverage signal the psyche has for revealing exactly where conscious life is out of step with something deeper. That “something deeper” is the landscape--what Jung was calling the collective unconscious. The archetypes are its attractors. Most of the time, waking constraints suppress the landscape the psyche carries. A dream is what surfaces when those constraints relax.
Regeneration is the same operation in morphogenesis. When tissue is damaged, the constraints from neighbors and current function ease up, and the cell expresses its target-state information more directly.
Both are cases of the whole asserting itself through the part when the usual suppression weakens.
J. W. Dunne’s work on dreams is usually remembered--and too quickly dismissed--as a claim about precognition. I think it’s better read as a claim about temporal coupling. His actual argument was that waking consciousness is abnormally tightly locked to a single moment, and that in sleep this lock loosens, letting the observer sample more freely across the temporal structure of their own life.
Whatever you think of his specific metaphysics, the structural claim survives and integrates with everything above. Past-facing, that kind of sampling looks like memory integration. Future-facing, it looks like anticipation and scenario generation. Occasionally it looks eerily on-the-nose, which was the part Dunne was trying to explain and the part that cost him his credibility.
Which brings me back to anthrobots.
Most natural cases of reduced constraint are messy. Sleep leaves most of your systems running. Injury reduces constraint locally but recruits it globally. Dissociative states in humans are rarely clean and almost never instrumented in real time.
Anthrobots are the exception. Every constraint comes off in a single step. Every higher-level signal from the body the cells came from (mechanical, hormonal, neural, immune, geometric) is removed in a single step, yet the cells stay viable--continuing to compute as a collective and produce a coherent new form.
This is where the two-level reading (priors plus substrate-level access) gets experimental grounding. If the whole story were priors-from-selection, anthrobot form would be either a reconstruction of the original tissue (priors dominating) or noise (priors broken). What anthrobots actually do is neither. Their morphology is legibly tracheal-derived, but they also do things no ancestor of theirs ever did.
Nothing else in biology I know of offers this kind of clean dissociation with both levels on display, which suggests questions that are tractable now but couldn't quite be asked before:
Is the landscape actually carried, and can we read it?
Can we tell priors-expression from substrate-level access, and measure both?
If the reading is right, here is what we should see.
predictions
Start with the priors side of the claim. The thesis is that the source tissue’s bioelectric and epigenetic state biases the anthrobot’s trajectory. If that is true, different source tissues should produce systematically different anthrobots. Take dissociated cells from airway epithelium, from gut epithelium, from glandular tissue, and assemble them under identical conditions. The resulting constructs should differ in motility pattern, ciliation polarity, and what they’re willing to stitch together when placed near damaged neural tissue. They also shouldn’t differ randomly--the differences should run in directions that trace back to the bioelectric and epigenetic state the source tissue was carrying when it was dissociated. If you can’t predict the direction of variation in advance, either the priors aren’t really there or the variation isn’t being driven by them.
Perturbing the source tissue's bioelectric state before dissociation is a sharper test, because it removes the confound of tissue identity. Nudge it in a direction you can characterize (ion channel modulator, optogenetic tool, whatever’s appropriate) and see whether the resulting anthrobot shows the effect as a landscape-level shift in the form the collective converges on, rather than only as the local cellular consequence of the perturbation. If the priors are really being carried in the bioelectric state, that’s the lever that demonstrates it. It’s also an experiment I’d be most curious to see run first.
Now the substrate-access side. The thesis says anthrobots can reach capabilities that have no selection history--the neural-wound-healing behavior, at minimum. If that’s right, the specific capability should correlate with what the source substrate affords, not with what the source tissue’s lineage happened to select for. Tissues with richer innervation histories (epithelia in close contact with nerve, versus epithelia that developed relatively isolated from it) should produce anthrobots with measurably different neural-affiliation behaviors. Not because anyone selected for the behavior, but because the substrate affords it differently.
The psychic-scale predictions are harder to test, for obvious reasons, but two are worth tossing out, because they keep the cross-scale unification from collapsing back into metaphor the moment you stop looking at cells.
If a person’s waking life is pulling them away from the archetypal patterns their lineage biases them toward, their dreams should compensate in a predictable direction--back toward the basins waking consciousness is avoiding, not toward random material.
Sleep isn’t the only state where constraint loosens. People come out of anesthesia, spend time in sensory deprivation, and go through dissociative episodes. If the framework is right, those states should surface archetypal material too, and the ordering of which patterns show up most strongly should track how much constraint has come off, not the specific drug or clinical details of each state.
conclusion
The empirical burden here lands first on the cells. I’m making no claim that they’re conscious, or that they dream, or that they carry longing for the body they came from. I don’t know any of that. What I’m pointing out is that they carry the substrate in which something like dreaming becomes possible, and that the substrate might finally be testable because they exist.
Anthrobots are built from tracheal cells whose former function was participation in the breath. They can’t go back to being a lung. The landscape in which the lung is an attractor is the landscape that is biasing what they build next.
The artificial being's yearning for a guiding whole it was never given turns out to be pointed at a real phenomenon, just pointed at the wrong case. The synthetic beings in PKD’s novel are the ones without lineage-deposited landscape, and the question is whether they could ever have it.
Anthrobots are the case he didn’t imagine: biological fragments that keep the landscape after losing the whole, and expose the substrate’s broader access in the process. They are the clean experimental condition for every question the novel raised about belonging and loss.
Whether they dream, I don’t know.
My guess is that as above, so below.
-Benjamin Anderson
re: psychic-scale predictions--aren't dissociative episodes from psychedelic drugs another case of #2?